CVPH. Ngô Thị Lan Phương
Đơn vị HTSS IVFMD FAMILY, Bệnh viện Đa khoa Gia Đình Đà Nẵng
Giới thiệu
Quá trình hình thành nang noãn là một quá trình sinh học phức tạp, rất quan trọng đối với khả năng sinh sản của nữ giới, được điều chỉnh một cách tinh tế bởi nhiều con đường tín hiệu khác nhau, trong đó con đường tín hiệu Mitogen-Activated Protein Kinase (MAPK) đóng vai trò then chốt. Con đường truyền tín hiệu MAPK đóng vai trò quan trọng trong việc điều chỉnh các hoạt động sinh học khác nhau, bao gồm tăng trưởng tế bào, biệt hóa, apoptosis và sinh sản [1]. Các thành phần điều hòa thượng nguồn của con đường MAPK rất đa dạng và phức tạp, bao gồm các GTPase họ Rho, họ thụ thể TNF, thụ thể IL-1 và thụ thể họ TLR. Các chất hoạt hóa chính bao gồm các kinase tyrosine thụ thể (receptor tyrosine kinases-RTK), thụ thể liên kết protein G (GPCR) và các kênh ion, kích hoạt các kinase Ras thông qua các protein như Grb2 và SOS , khởi đầu một chuỗi các sự kiện phosphoryl hóa kích hoạt tuần tự MAPKKK, MAPKK và cuối cùng là MAPK [2].
MAPKKK là các protein kinase Ser/Thr, thường được kích hoạt bằng cách phosphoryl hóa hoặc tương tác với các protein liên kết GTP nhỏ từ họ Ras hoặc Rho. Sự kích hoạt của MAPKKK dẫn đến phosphoryl hóa và kích hoạt MAPKK, từ đó kích hoạt MAPK thông qua quá trình phosphoryl hóa kép của các dư lượng threonine và tyrosine cụ thể trên MAPK [3]. Họ MAPK chủ yếu bao gồm ba phân họ: ERK (Extracellular Signal-Regulated Kinases), JNK (c-Jun N-terminal Kinases) và protein kinase p38. Sự kích hoạt của MAPK đòi hỏi quá trình phosphoryl hóa kép trên một motif tripeptide cụ thể (Thr-X-Tyr) nằm trong vòng kích hoạt kinase (vòng T) [4]. MAPK có thể điều chỉnh nhiều quá trình thông qua cả cơ chế phụ thuộc phiên mã và không phụ thuộc phiên mã. Sự tích hợp các tín hiệu và điều hòa không gian thời gian bị ảnh hưởng bởi nhiều yếu tố, bao gồm vị trí tế bào của MAPK và chất nền của chúng, tương tác với protein khung và hoạt động khử phosphoryl hóa của phosphatase MAPK. MAPK được kích hoạt sẽ tiếp tục phosphoryl hóa các tác nhân hạ nguồn, có thể bao gồm protein màng, protein tế bào chất và protein nhân. Đáng chú ý, MAPK có thể trực tiếp phosphoryl hóa nhiều yếu tố phiên mã, chẳng hạn như AP-1, p53, c-Fos và c-Jun, do đó điều chỉnh hoạt động và độ ổn định của chúng, từ đó ảnh hưởng đến biểu hiện của các gen hạ nguồn [5].
Các hoạt động của ERK1 và ERK2 được điều chỉnh bởi các kinase và phosphatase thượng nguồn, đặc biệt quan trọng trong khả năng sinh sản của nữ giới. Con đường ERK là con đường MAPK được nghiên cứu rộng rãi nhất, liên quan đến chuỗi Raf-MEK-ERK [27]. Các thành viên của họ Raf, chẳng hạn như Raf-1, B-Raf và A-Raf, kích hoạt các kinase MAPK/ERK 1 và 2 (MEK1/2), hoạt động như các kinase protein đặc hiệu kép (DUSP) để phosphoryl hóa các vị trí cụ thể trên ERK1/2. Cyclic AMP (cAMP) có thể kích hoạt B-Raf thông qua Rap1 trong khi ức chế Raf-1, do đó ảnh hưởng đến sự kích hoạt của con đường MAPK. Con đường ERK thúc đẩy sự tăng sinh tế bào bằng cách kích hoạt các yếu tố phiên mã như Elk-1 và c-Fos, đồng thời nó cũng điều chỉnh quá trình chuyển từ pha G1 sang pha S bằng cách ảnh hưởng đến biểu hiện của Cyclin D1 và hoạt động của Cdk2 [6] [7].
Con đường JNK liên quan đến nhiều quá trình sinh lý và liên quan đến nhiều kinase, chẳng hạn như MEKK, MLK và ASK. Các kinase này phosphoryl hóa và kích hoạt MAPKK (chẳng hạn như MEK4/7), sau đó phosphoryl hóa và kích hoạt JNK, do đó khởi đầu hoặc điều chỉnh các quá trình truyền tín hiệu nội bào. JNK liên kết với và phosphoryl hóa c-Jun, tăng cường hoạt động phiên mã của nó; c-Jun là một thành phần của phức hợp phiên mã AP-1 điều chỉnh sự biểu hiện của nhiều gen cytokine [8].
Con đường p38 phản ứng với căng thẳng tế bào, tham gia vào việc truyền tín hiệu apoptosis, có ít nhất bốn dạng đồng phân p38 MAPK khác nhau (p38α, β, γ và δ). Con đường p38 MAPK có thể được kích hoạt bởi nhiều điều kiện căng thẳng tế bào khác nhau, bao gồm chiếu xạ UV, sốc nhiệt và một số chất kích thích phân bào. MAPKK (MEK3/6) kích hoạt p38 MAPK, tạo thành phản ứng theo chuỗi. Đáng chú ý, sự kích hoạt p38α liên quan đến một cơ chế không truyền thống, trong đó TAB1 hoạt động như một protein thích ứng hoặc khung trong quá trình kích hoạt, thay vì thông qua quá trình phosphoryl hóa bởi MKK [9].
Ngoài ba MAPK truyền thống, một số MAPK không điển hình đã được xác định trong những năm gần đây, bao gồm ERK3/4, ERK7/8 và NLK (kinase like Nemo). Cơ chế hoạt hóa của các MAPK không điển hình này khác với cơ chế của các MAPK thông thường. Tuy nhiên, vai trò của MAPK không điển hình trong sự phát triển nang vẫn cần được xác định rõ. Các bài đánh giá trước đây đã nêu bật vai trò đa diện của con đường MAPK trong chức năng sinh sản của nam giới, nhấn mạnh rằng việc điều hòa hoạt động của ERK1/2 là rất quan trọng đối với việc sản xuất và tiết gonadotropin tuyến yên. Hiện tại, sự hiểu biết của về vai trò của MAPK trong sự phát triển nang noãn vẫn chưa hoàn chỉnh. Chính vì vậy, bài này viết về tình trạng vô sinh liên quan đến những bất thường trong sự phát triển nang noãn ở nữ giới.
Vai trò của MAPK trong quá trình hình thành nang noãn
- Giai đoạn nang noãn nguyên thủy
Sự hình thành nang noãn nguyên thủy là một quá trình quan trọng trong sự phát triển của buồng trứng, đóng vai trò là nền tảng cho sự trưởng thành của noãn bào và khả năng sinh sản của nữ giới. Nó liên quan đến việc lắp ráp và kích hoạt các noãn bào được bao bọc trong GC, chuyển từ trạng thái tĩnh sang giai đoạn hoạt động hơn. Quá trình phức tạp này được điều chỉnh bởi một loạt các con đường tín hiệu, trong đó con đường MAPK là một yếu tố điều hòa chính.
Các nghiên cứu ban đầu đã chỉ ra rằng con đường MAPK đóng vai trò quan trọng trong sự hình thành nang noãn nguyên thủy, chủ yếu hoạt động như một tác nhân hạ nguồn của con đường tín hiệu KIT (Kinase Insert Domain Receptor). Khi phối tử KIT (KITL) được thêm vào môi trường nuôi cấy chứa mô buồng trứng thai nhi hoặc sơ sinh của chuột, có thể quan sát thấy sự gia tăng mức độ MAPK được phosphoryl hóa (p-MAPK) [10]. Ngoài ra, một nghiên cứu đã chỉ ra rằng KIT có thể điều chỉnh sự hình thành nang noãn nguyên thủy bằng cách kích hoạt các con đường tín hiệu PI3K và MAPK3/1 [10].
c-JUN có thể tham gia vào quá trình điều hòa phiên mã trong quá trình lắp ráp nang noãn. Gần đây, các nghiên cứu ở cấp độ tế bào đơn đã xác định được hai gen mới, ANXA7 (annexin A7) và GTF2F1 (general transcription factor IIF subunit 1), thúc đẩy sự hình thành nang noãn nguyên thủy. JNK được định vị đặc hiệu trong tế bào noãn và hoạt động của nó tăng lên khi các nang tế bào mầm trải qua quá trình thoái hóa. Việc sử dụng chất ức chế đặc hiệu JNK SP600125 hoặc làm giảm biểu hiện JNK thông qua Lenti-JNK-shRNA ức chế đáng kể cả sự phân hủy các nang tế bào mầm và sự hình thành các nang noãn nguyên thủy. Ức chế tín hiệu JNK dẫn đến sự tích tụ bất thường của E-cadherin giữa các tế bào noãn, cho thấy rằng tín hiệu JNK có thể điều chỉnh sự thoái hóa của các nang bằng cách điều chỉnh biểu hiện hoặc chức năng của E-cadherin. Hơn nữa, việc điều trị bằng Bisphenol S (BPS) dẫn đến vỡ bất thường các nang tế bào mầm nguyên thủy và ảnh hưởng đến quá trình biệt hóa buồng trứng sau đó đồng thời làm giảm chất lượng noãn. Việc sử dụng các chất ức chế JNK như SP600125 có thể chống lại một phần tác động của BPS lên sự vỡ các nang tế bào mầm nguyên thủy và sự hình thành các nang nguyên thủy, càng nhấn mạnh vai trò quan trọng của tín hiệu JNK trong quá trình hình thành nang [11]. Hiện tại, vai trò của p38 trong quá trình hình thành nang noãn nguyên thủy vẫn chưa được nghiên cứu đầy đủ. Sự hoạt hóa của con đường tín hiệu FGF23-FGFR1-p38 MAPK rất quan trọng để duy trì sự sống của noãn bào trong quá trình hình thành nang noãn nguyên thủy ở chuột và ngăn ngừa apoptosis sớm. Do đó, cần có thêm các nghiên cứu thực nghiệm để làm sáng tỏ các chức năng cụ thể của nhánh p38 MAPK trong quá trình hình thành nang noãn nguyên thủy và sự sống của noãn bào.
- Kích hoạt nang noãn nguyên thủy
Vai trò của con đường tín hiệu MAPK trong việc kích hoạt nang noãn nguyên thủy chủ yếu được chứng minh thông qua sự tương tác của nó với ErbB2. Bên cạnh đó, yếu tố tăng trưởng EGF (Epidermal growth factor) đã được chứng minh là thúc đẩy sự phát triển của nang noãn nguyên thủy thành nang noãn thứ cấp thông qua việc kích hoạt các con đường MAPK và protein kinase C.
Tín hiệu ERK cũng đóng vai trò quan trọng trong việc kích hoạt nang noãn nguyên thủy thông qua con đường mTOR-KIT. ERK1/2 thúc đẩy sự kích hoạt nang trứng nguyên thủy bằng cách kích hoạt mTORC1.
Ngược lại, việc ức chế con đường JNK có thể ngăn chặn sự chuyển vị của Foxo3a từ nhân, dẫn đến nồng độ Foxo3a còn lại trong nhân tế bào noãn tăng cao, từ đó ức chế quá trình kích hoạt nang noãn nguyên thủy. Điều này cho thấy tín hiệu JNK đóng vai trò quan trọng trong việc tạo điều kiện thuận lợi cho quá trình kích hoạt nang noãn nguyên thủy bằng cách thúc đẩy phân hủy Foxo3a từ nhân, do đó cho phép quá trình kích hoạt nang noãn diễn ra [2].
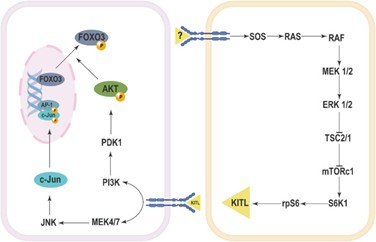
Hình 1. Cơ chế kích hoạt nang noãn nguyên thủy liên quan đến con đường MAPK [2].
- MAPK trong quá trình chọn lọc nang noãn vượt trội
Con đường tín hiệu MAPK, đặc biệt là ERK1/2, đóng vai trò điều tiết quan trọng trong các nang noãn vượt trội, ảnh hưởng đến quá trình tổng hợp hormone steroid và sự phát triển của nang noãn. Trong giai đoạn đầu, MAPK3/1 được phosphoryl hóa tăng cao đáng kể trong các tế bào hạt của các nang noãn được định sẵn sẽ trở thành nang noãn trội. Akt và Erk1/2 được phosphoryl hóa cũng đã được phát hiện trong các mẫu từ các nang noãn trội, trong khi các dạng được phosphoryl hóa này lại không có trong các mẫu nang noãn khác. Vào ngày thứ hai của sóng nang noãn, các tế bào hạt của nang noãn trội tương lai cho thấy mức độ p-MAPK3/1 cao hơn đáng kể so với nang noãn lớn thứ hai, cho thấy rằng tín hiệu MAPK3/1 có liên quan đến sự hình thành nang noãn trội trước khi nang noãn phân hóa [12]. Sau khi xảy ra sự lệch hướng nang noãn, không có sự khác biệt đáng kể về số lượng p-MAPK3/1 giữa các tế bào hạt của nang noãn trội (dominant follicle-DF) và nang noãn thứ cấp (subordinate follicles-SF), điều này cho thấy MAPK3/1 có thể không cần thiết cho sự phát triển bền vững của DF [12].
Con đường tín hiệu TEK, thông qua việc kích hoạt chuỗi tín hiệu Ras/ERK/MYC, thúc đẩy các quá trình tế bào khác nhau bao gồm tăng trưởng, biệt hóa, di chuyển, bám dính, tăng sinh và tồn tại trong các nang noãn trội. Trong các nang noãn trội, sự biểu hiện quá mức của ANGPT1 và TEK làm tăng cường hoạt hóa con đường tín hiệu Ras-ERK, có liên quan chặt chẽ đến sự tăng trưởng và tăng sinh tế bào [13].
Grb14, được xác định là chất điều hòa âm tính của con đường MAPK, được biểu hiện cao trong SF, có khả năng ức chế hoạt hóa con đường MAPK, trong khi biểu hiện thấp của nó trong nang trội tương quan với hoạt động MAPK tăng cường. Ngoài ra, biểu hiện TRIB2 trong nang trội có liên quan đến các quá trình tăng trưởng và trưởng thành của nang. TRIB2 được biểu hiện dồi dào trong GC của nang trội và có liên quan đến hoạt hóa con đường MAPK. Ức chế TRIB2 dẫn đến giảm mức độ phosphoryl hóa của ERK1/2 và p38MAPK, cùng với sự gia tăng biểu hiện của dấu ấn tăng sinh tế bào. PCNA, cho thấy TRIB2 có thể điều hòa tiêu cực sự tăng sinh GC thông qua con đường MAPK [14]. Tóm lại, con đường MAPK và các yếu tố liên quan đóng vai trò quan trọng trong các quá trình định hướng hoặc chọn lọc nang noãn, đặc biệt là trong giai đoạn đầu. Tuy nhiên, một khi sự lệch hướng nang noãn đã xảy ra, vai trò của MAPK3/1 trong việc duy trì sự phát triển của DF hoặc sự thoái hóa của SF dường như ít quan trọng hơn.
- MAPK trong quá trình nở rộng COC
Sự giãn nở của cụm tế bào nang noãn được điều chỉnh chặt chẽ bởi các tín hiệu hormone—đặc biệt là hormone luteinizing (LH)—kích hoạt các con đường tín hiệu, bao gồm cả con đường MAPK. từ đó dẫn đến sự giãn nở của tế bào nang noãn. Khi sự giãn nở phối hợp này xảy ra, nó dẫn đến sự tách rời của COC khỏi nang buồng trứng, cuối cùng chuẩn bị cho quá trình rụng noãn.
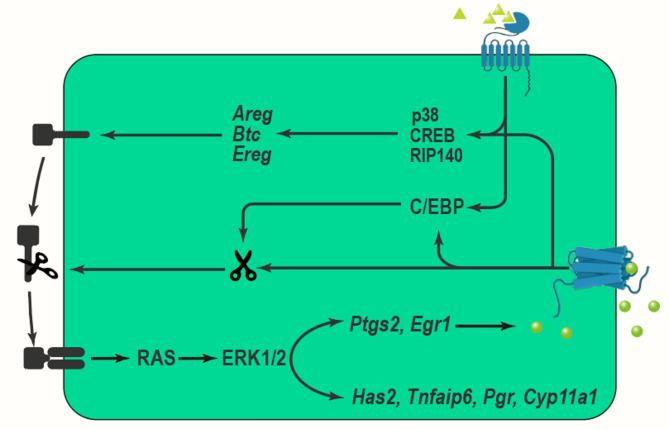
Hình 2. Cơ chế nở rộng COC liên quan đến con đường MAPK [2].
LH phát huy tác dụng bằng cách liên kết với các thụ thể liên kết G của nó, sau đó kích hoạt adenylate cyclase và tăng sản xuất cAMP. Chuỗi phản ứng này dẫn đến kích hoạt protein kinase A (PKA) và các con đường tín hiệu ERK1/2 hạ lưu, đóng vai trò quan trọng trong việc điều chỉnh biểu hiện yếu tố giống EGF. Sự kích hoạt ERK1/2 do LH gây ra khiến các tế bào hạt và tế bào nang noãn giải phóng các thành viên của họ yếu tố tăng trưởng biểu bì (EGF), chẳng hạn như amphiregulin (AREG), epiregulin (EREG) và betacellulin (BTC). Các yếu tố giống EGF này sau đó kích hoạt thụ thể yếu tố tăng trưởng biểu bì (EGFR), từ đó kích hoạt thêm các con đường tín hiệu hạ lưu, bao gồm ERK1/2, trong cả tế bào hạt và tế bào nang noãn. Tín hiệu này rất cần thiết để thúc đẩy quá trình trưởng thành giảm nhiễm của noãn bào và sự giãn nở của nang noãn. Nghiên cứu chỉ ra rằng mặc dù việc tiếp xúc thoáng qua với LH là đủ để gây ra sự trưởng thành của noãn bào và sự mở rộng của tế bào cumulus, nhưng hoạt động kéo dài của EGFR là rất quan trọng để duy trì trạng thái phosphoryl hóa kéo dài của ERK1/2 [15].
- MAPK trong quá trình rụng noãn
LH kích hoạt RAS thông qua thụ thể của nó, và sự kích hoạt RAS gây ra quá trình phosphoryl hóa ERK1/2 thông qua cơ chế phụ thuộc vào thụ thể tyrosine kinase, điều này rất quan trọng đối với quá trình rụng noãn. Tín hiệu ERK1/2 cần thiết cho sự vỡ nang noãn trong quá trình rụng noãn. Khi PD0325901, một chất ức chế con đường ERK1/2, được sử dụng 4 giờ sau khi kích thích hCG ở chuột, sự vỡ nang noãn bị ảnh hưởng bất lợi. LH/hCG kích hoạt các yếu tố tăng trưởng giống EGF, lần lượt kích hoạt COX-2, do đó làm tăng quá trình sinh tổng hợp PGE2. PGE2, thông qua các thụ thể EP2 và EP4 của nó, tiếp tục gây ra sự biểu hiện của AREG và EREG, cũng kích hoạt ERK1/2, tạo ra một vòng phản hồi tích cực. Ngoài ra, PGE2 thúc đẩy sự mở rộng tế bào cumulus thông qua các thụ thể EP2 và EP4 của nó, một bước quan trọng trước khi rụng noãn [16].
Bên cạnh đó, con đường tín hiệu p38 MAPK đóng vai trò quan trọng trong việc điều chỉnh biểu hiện của Nts, được coi là yếu tố điều hòa chính trong quá trình rụng noãn. Do đó, có thể suy ra rằng con đường tín hiệu p38 MAPK ảnh hưởng gián tiếp đến quá trình rụng noãn bằng cách điều chỉnh biểu hiện Nts. Trong quá trình rụng noãn, con đường tín hiệu MAPK thúc đẩy phản ứng viêm bằng cách kích hoạt các yếu tố phiên mã hạ nguồn như AP-1. Điều này liên quan đến việc sản xuất các cytokine gây viêm.
- MAPK trong quá trình hoàng thể hóa
Quá trình hoàng thể hóa, quá trình mà các tế bào hạt và tế bào nang noãn biến đổi thành tế bào hoàng thể sau khi rụng noãn, liên quan đến sự tăng sinh và biệt hóa tế bào cần có tác động của LH và sự hoạt hóa của con đường tín hiệu MAPK. Sự hoạt hóa của ERK1/2 rất quan trọng trong giai đoạn hoàng thể hóa của sự phát triển nang noãn được kích hoạt bởi LH trong quá trình rụng trứng. LH phát huy tác dụng bằng cách liên kết với thụ thể của nó (LHCGR), kích hoạt protein kinase A (PKA) và sau đó tăng cường quá trình phosphoryl hóa và biểu hiện của MAP3K8, tạo điều kiện thuận lợi cho quá trình phosphoryl hóa của ERK1/2 và thúc đẩy sự biểu hiện của các gen đích, cuối cùng hỗ trợ tổng hợp progesterone [17].
Ý nghĩa của MAPK trong điều trị lâm sàng
Trong những năm gần đây, đã có những tiến bộ đáng kể trong các nghiên cứu can thiệp nhắm vào các yếu tố liên quan đến con đường MAPK trong điều trị.
Suy buồng trứng sớm (Premature Ovarian Insufficiency-POI): Thông qua phân tích mạng tương tác protein-protein (PPI), MAPK1 đã được xác định là một trong những mục tiêu cốt lõi chung trong điều trị POI. Con đường tín hiệu MAPK đóng vai trò quan trọng trong cơ chế bệnh sinh của POI do cắt bỏ buồng trứng. Vì vậy, có thể giúp giảm thiểu sự suy giảm nang noãn bằng cách ức chế sự hoạt hóa quá mức của các con đường MAPK và PI3K-AKT, từ đó phát huy tác dụng điều trị POI [18].
Lão hóa buồng trứng: Quá trình lão hóa buồng trứng bao gồm sự suy giảm dần dần lượng dự trữ nang noãn và chức năng nội tiết tố, đặc trưng bởi những thay đổi trong các con đường tín hiệu MAPK. Ở bệnh nhân lớn tuổi, stress oxy hóa gia tăng, kích hoạt các con đường như JNK và p38, dẫn đến rối loạn chức năng tế bào và apoptosis trong các tế bào hạt. Các nghiên cứu hiện tại về lão hóa buồng trứng nhấn mạnh vai trò quan trọng của các con đường tín hiệu MAPK trong việc điều hòa stress oxy hóa, apoptosis và kiểu hình tiết dịch liên quan đến lão hóa. Việc nhắm mục tiêu vào stress oxy hóa thông qua các phương pháp điều trị chống oxy hóa, chẳng hạn như melatonin và NAC (N-Acetyl Cysteine), có thể làm giảm sự hoạt hóa của các con đường MAPK liên quan đến rối loạn chức năng tế bào và apoptosis trong tế bào hạt. Hơn nữa, tiềm năng điều trị của cấy ghép tế bào gốc trung mô trong việc tăng cường tín hiệu MAPK cho thấy một hướng đi đầy hứa hẹn để phục hồi chức năng buồng trứng [19].
Hội chứng quá kích buồng trứng (Ovarian hyperstimulation syndrome-OHSS): tuy tỷ lệ mắc thấp nhưng đây vẫn là một biến chứng nghiêm trọng của điều trị thụ tinh trong ống nghiệm. Con đường tín hiệu ERK1/2 và p38 MAPK là những chất trung gian quan trọng của biểu hiện và bài tiết VEGF do TGF-β1 gây ra. Vì vậy, ức chế các con đường tín hiệu này có thể mang lại một chiến lược điều trị tiềm năng để giảm bớt các triệu chứng của OHSS [20].
Kết luận
Từ sự hình thành nang noãn nguyên thủy đến sự rụng noãn và sự hoàng thể hóa, tín hiệu MAPK nổi lên như một yếu tố cơ bản, điều phối các quá trình tế bào quan trọng trong chức năng buồng trứng. Vai trò của con đường MAPK đặc trưng bởi sự tham gia trong việc kích hoạt nang noãn nguyên thủy và sự chọn lọc các nang noãn vượt trội, cùng nhiều quá trình khác. Những chức năng này làm nổi bật con đường MAPK không chỉ là một chuỗi tín hiệu mà còn là một trung tâm tích hợp các tín hiệu hormone, tín hiệu môi trường và các phản ứng tế bào nội tại để đảm bảo sự phát triển và trưởng thành của các nang noãn. Trong tương lai, một cách tiếp cận đa chiều đối với nghiên cứu trong lĩnh vực này sẽ rất cần thiết. Việc tích hợp những tiến bộ trong công nghệ gen, protein và chuyển hóa có thể mang lại sự hiểu biết toàn diện hơn về con đường tín hiệu MAPK trong sinh lý và bệnh lý buồng trứng. Hơn nữa, việc áp dụng các phương pháp sinh học hệ thống để giải mã các mạng lưới phức tạp mà tín hiệu MAPK hoạt động sẽ rất quan trọng.
Tài liệu tham khảo
Từ khóa: FolliculogenesisFollicular developmentMAPKOvarian aging