Nội dung chính
- 1.Cơ chế sửa chữa DNA bị bất hoạt ở tinh trùng trưởng thành
- 2. Các con đường sửa chữa DNA sau thụ tinh
- 2.1. Sửa chữa tổn thương sợi đơn DNA
- 2. Sửa chữa tổn thương sợi đôi DNA
- 3. Ngưỡng sửa chữa của noãn
- 4. Ý nghĩa trong thực hành lâm sàng
- 5. Những hạn chế và thách thức hiện tại
- 6. Định hướng cho tương lai
- Tài liệu tham khảo
Quá trình sinh tinh là một chuỗi các sự kiện biệt hóa phức tạp nhằm tạo ra tinh trùng có khả năng di động và thụ tinh. Giai đoạn cuối của quá trình này đặc trưng bởi việc thay thế phần lớn các protein Histone bằng các protein Transition Nuclear Proteins (TNP) và cuối cùng là Protamine (Balder và cs. 2024) (Hình 1). Sự tái cấu trúc chất nhiễm sắc này dẫn đến sự nén chặt nhân tinh trùng gấp 6 đến 10 lần so với tế bào soma, tạo ra cấu trúc dạng Toroid giúp bảo DNA khỏi các tác nhân gây hại vật lý và hóa học (Anais và cs. 2019, Balder và cs. 2024). Tuy nhiên, đặc điểm cấu trúc này cũng làm tinh trùng trưởng thành bị bất hoạt về mặt phiên mã và dịch mã, dẫn đến sự thiếu hụt các enzyme cần thiết cho việc tự sửa chữa DNA (Aitken và cs. 2023, Li và cs. 2025).
Tổn thương DNA tinh trùng, được định nghĩa qua chỉ số đứt gãy DNA tinh trùng (Sperm DNA Fragmentation-SDF), phát sinh từ ba cơ chế chính: đóng gói nhiễm sắc chất không hoàn chỉnh, stress oxy hóa và quá trình chết tế bào theo chương trình bị trì hoãn (Aitken và cs. 2023, Pardiñas và cs. 2026). Theo giả thuyết hai bước, bước đầu tiên liên quan đến việc đóng gói kém chất nhiễm sắc trong tinh hoàn làm lộ ra các vùng DNA nhạy cảm (Aitken và cs. 2023). Bước thứ hai diễn ra khi các vùng này bị tấn công bởi các gốc tự do ROS, hoặc các enzyme nuclease trong quá trình di chuyển qua mào tinh (Aitken và cs. 2023, Esteves 2025).
Một đặc điểm cấu trúc độc đáo của tinh trùng là nhân và ty thể nằm ở các khoang tế bào khác nhau (Aitken và cs. 2023). Trong các tế bào soma, quá trình apoptosis giải phóng các enzyme nuclease vào nhân để phân hủy DNA, nhưng ở tinh trùng, cấu trúc tế bào ngăn cản các enzyme này tiếp cận khoang chứa nhân (Aitken và cs. 2023, Gunes và cs. 2018). Ngược lại, các ROS như hydro peroxide (H2O2) không mang điện tích nên có thể xuyên qua màng ngăn để tấn công các vùng DNA không được nén chặt bởi Protamine (Aitken và cs. 2023). Các tổn thương oxy hóa phổ biến nhất là sự hình thành dẫn xuất 8-hydroxy-2'-deoxyguanosine (8-OHdG) (Pardiñas và cs. 2026).
Mặc dù tinh trùng người chứa enzyme 8-oxoguanine DNA glycosylase 1 (OGG1) để nhận diện và cắt bỏ base bị oxy hóa, nhưng hệ thống Base Excision Repair (BER - Sửa chữa cắt bỏ gốc) bị khiếm khuyết do thiếu các thành phần hỗ trợ như Apurinic/apyrimidinic endonuclease 1 (APE1) và X-ray repair cross-complementing protein 1 (XRCC1) (Aitken và cs. 2023, Li và cs. 2025). Hệ quả là các base hỏng và các vị trí abasic không được xử lý sẽ được chuyển trực tiếp vào noãn tại thời điểm thụ tinh (Pardiñas và cs. 2026). Do đó, sự phát triển của phôi phụ thuộc hoàn toàn vào khả năng noãn trong việc nhận diện và sửa chữa các tổn thương này trước khi quá trình nhân đôi DNA bắt đầu (Newman và cs. 2022).
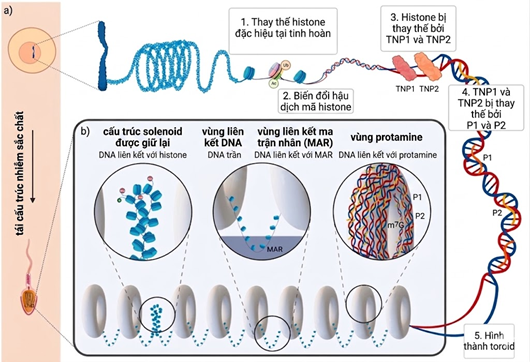
Hình 1. Quá trình tái cấu trúc và tổ chức chất nhiễm sắc của tinh trùng. Quá trình này bắt đầu bằng việc thay thế các Histone bằng các biến thể đặc hiệu, sau đó trải qua các biến đổi hậu dịch mã để các protein chuyển tiếp (TNP) và cuối cùng là Protamine (P1, P2) đến thay thế, giúp bộ gen tinh trùng đạt trạng thái siêu nén chặt. Cấu trúc nhân tinh trùng trưởng thành gồm ba thành phần: vùng DNA liên kết Histone (Solenoid) được giữ lại, vùng gắn kết khung nhân (MAR) và các khối Toroid nén chặt do DNA liên kết với Protamine. (Nguồn Balder và cs. 2024)
Sau khi thụ tinh, nhân tinh trùng trải qua quá trình giải nén nhiễm sắc chất để tạo điều kiện cho các yếu tố báo hiệu và enzyme sửa chữa của noãn tiếp cận bộ gen tinh trùng (Newman và cs. 2022). Vì noãn và phôi giai đoạn sớm bị trì hoãn phiên mã cho đến khi quá trình hoạt hóa bộ gen phôi diễn ra, mọi hoạt động sửa chữa đều phụ thuộc vào kho dự trữ mRNA và các protein được tổng hợp trong quá trình trưởng thành nang noãn (Newman và cs. 2022, Pardiñas và cs. 2026).
Hệ thống Base Excision Repair (BER - Sửa chữa cắt bỏ gốc) là con đường chủ đạo để xử lý các base bị oxy hóa, đặc biệt là các tổn thương mutagenic như 8-hydroxy-2'-deoxyguanosine (8-OHdG) (García-Rodríguez và cs. 2018, Pardiñas và cs. 2026). Một đặc điểm quan trọng trong giai đoạn này là sự hợp tác sinh học giữa hai giao tử: tinh trùng mang theo enzyme 8-Oxoguanine DNA Glycosylase 1 (OGG1) để nhận diện và loại bỏ base hỏng, tạo ra vị trí không chứa base. Tuy nhiên, do tinh trùng thiếu hụt các protein kế cận, quá trình này chỉ được hoàn tất bởi các enzyme Apurinic/apyrimidinic Endonuclease 1 (APE1) và X-ray Repair Cross-Complementing Protein 1 (XRCC1) có nồng độ cao trong bào tương noãn (Lord và Aitken 2015, Pardiñas và cs. 2026) (Hình 2).
Đối với các đứt gãy sợi đơn trực tiếp, hệ thống Sửa chữa đứt gãy sợi đơn (Single-Strand Break Repair-SSBR) được kích hoạt thông qua protein Poly (ADP-Ribose) Polymerase 1 (PARP1). Protein này đóng vai trò là cảm biến liên kết với vị trí đứt gãy, huy động phức hợp sửa chữa bao gồm DNA Polymerase Beta và DNA Ligase III để lấp đầy khoảng trống và gắn nối sợi DNA (García-Rodríguez và cs. 2018, Pardiñas và cs. 2026) (Hình 2).
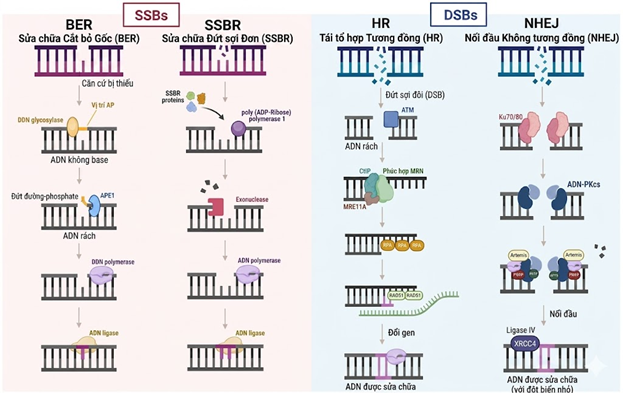
Hình 2. Các cơ chế của noãn sửa chữa đứt gãy DNA tinh trùng. Trình tự các bước phân tử trong bốn lộ trình chính: sửa chữa cắt bỏ base (BER) và sửa chữa đứt gãy sợi đơn (SSBR) đối với các tổn thương sợi đơn (SSBs); tái tổ hợp tương đồng (HR) và nối đầu không tương đồng (NHEJ) đối với các tổn thương sợi đôi (DSBs). Trình tự phản ứng bắt đầu từ giai đoạn nhận diện tổn thương bởi các protein cảm biến (như Enzyme glycosylase, ATM hoặc phức hợp Ku70/80), tiếp theo là các bước cắt và xử lý đầu DNA (như vị trí AP, phức hợp MRN hoặc RPA), và kết thúc bằng việc tổng hợp bù đắp bởi DNA polymerase cùng quá trình nối kín sợi bởi DNA ligase để phục hồi tính toàn vẹn của bộ gen.
(SSB/DSB: Đứt gãy sợi đơn/sợi đôi; BER/SSBR: Sửa chữa cắt bỏ base/đứt gãy sợi đơn; HR/NHEJ: Tái tổ hợp tương đồng/Nối đầu không tương đồng; AP: Vị trí abasic; MRN: Phức hợp Mre11-Rad50-Nbs1) (Nguồn: Pardinas và cs. 2026)
Đứt gãy sợi đôi là loại tổn thương nghiêm trọng nhất đến tính ổn định của bộ gen phôi (Alvarez và cs. 2023). Hợp tử triển khai hai con đường chính với vai trò bổ trợ lẫn nhau tùy theo giai đoạn chu kỳ tế bào:
Tái tổ hợp tương đồng (Homologous Recombination-HR) là con đường sửa chữa có độ chính xác cao nhất vì sử dụng sợi chị em làm khuôn mẫu. Quá trình này bắt đầu khi cảm biến Ataxia Telangiectasia Mutated (ATM) nhận diện tổn thương và huy động phức hợp Mre11-Rad50-Nbs1 (MRN) cùng protein DNA Repair Protein RAD51 Homolog 1 (RAD51) (Pardiñas và cs. 2026, Li và cs. 2025). Con đường HR hoạt động mạnh mẽ nhất ở giai đoạn S và G2 của chu kỳ hợp tử giúp ngăn chặn sự cố định các đột biến vào bộ gen phôi (Alvarez và cs. 2023) (Hình 2).
Nối đầu không tương đồng (Non-Homologous End Joining-NHEJ) là một con đường khác. Khác với HR, con đường này không cần khuôn mẫu nên diễn ra nhanh chóng nhưng dễ gây sai sót di truyền như mất đoạn hoặc chèn đoạn. Phức hợp protein X-ray Repair Cross-Complementing Protein 6 (Ku70) và X-ray Repair Cross-Complementing Protein 5 (Ku80) sẽ liên kết trực tiếp với các đầu đứt gãy, huy động DNA-Dependent Protein Kinase Catalytic Subunit (DNA-PKcs) và DNA Ligase IV để nối lại các đầu DNA (Li và cs. 2025, Pardiñas và cs. 2026). NHEJ là cơ chế sửa chữa chiếm ưu thế trong giai đoạn G1 của hợp tử trước khi quá trình nhân đôi DNA diễn ra (Alvarez và cs. 2023) (Hình 2,3).
Sự chuyển đổi linh hoạt giữa các con đường này đảm bảo rằng phôi có thể xử lý các tổn thương di truyền từ tinh trùng trong một khung thời gian có hạn trước khi bước vào các đợt phân bào liên tiếp (Newman và cs. 2022).
a. Trước EGA
Khả năng của noãn trong việc bù trừ các tổn thương di truyền từ tinh trùng có giới hạn định lượng nhất định. Juan và cs. (2023) đã đề xuất sự tồn tại của một ngưỡng không thể quay đầu (Threshold of no return). Theo một số nghiên cứu, noãn có khả năng sửa chữa hiệu quả các tổn thương DNA tinh trùng khi mức độ tổn thương dưới ngưỡng 8%.
Khi mức độ SDF vượt quá khả năng xử lý của noãn, phôi sẽ không thể phát triển đến các mốc quan trọng. Mức độ đứt gãy DNA càng cao và càng phức tạp (đặc biệt là đứt gãy sợi đôi) gây áp lực lên hệ thống sửa sai của noãn càng lớn dẫn đến việc không thể hoàn thành quá trình sửa chữa trước khi bắt đầu các đợt phân bào.
b. Sau EGA
Một đặc điểm quan trọng của tổn thương DNA tinh trùng là thường không ngăn cản quá trình thụ tinh hay sự phân chia ban đầu của phôi. Điều này là do phôi giai đoạn sớm được điều phối phát triển chủ yếu bởi dự trữ protein và mRNA có sẵn của noãn (Hình 3).
Tuy nhiên, khi quá trình EGA diễn ra ở giai đoạn phôi từ 4 đến 8 tế bào, các tổn thương DNA tinh trùng không được sửa chữa trước đó sẽ kích hoạt các điểm kiểm soát chu kỳ tế bào, dẫn đến hiện tượng chậm phân chia, ngưng phát triển phôi hoặc sẩy thai (Hình 3), hiện tượng này được gọi là Late Paternal Effect.
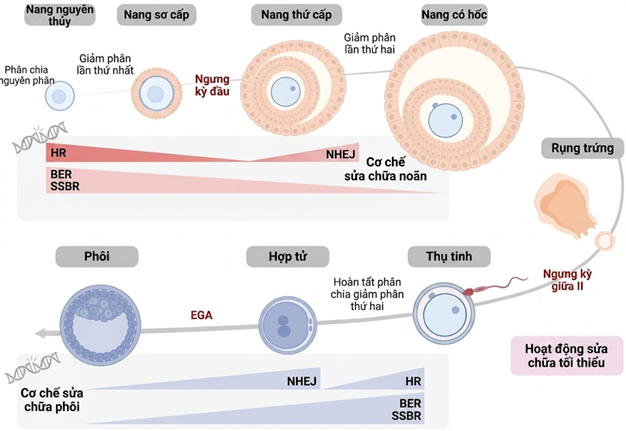
Hình 3. Các cơ chế sửa chữa DNA của noãn và phôi qua các giai đoạn phát triển nang trứng và phát triển phôi. Sự thay đổi hoạt tính của các lộ trình sửa chữa từ khi nang trứng hình thành cho đến giai đoạn phôi. Trong quá trình trưởng thành của noãn, tế bào chủ yếu dựa vào tái tổ hợp tương đồng (HR) để sửa chữa các đứt gãy sợi đôi (DSB), bên cạnh hoạt động song hành của lộ trình sửa chữa cắt bỏ gốc (BER) và sửa chữa đứt gãy sợi đơn (SSBR). Khi tiến gần đến giai đoạn rụng trứng, do sự thiếu hụt các nhiễm sắc tử chị em, hoạt tính HR giảm dần và nối đầu không tương đồng (NHEJ) trở thành cơ chế chủ đạo để xử lý DSB. Sau khi rụng trứng, hoạt động sửa chữa bị giảm thiểu đáng kể nhưng sẽ được tái kích hoạt ngay sau khi thụ tinh. Nếu DNA tinh trùng bị tổn thương, noãn sẽ khởi động các hệ thống HR, NHEJ, BER và SSBR để khắc phục sai sót, đảm bảo sự kích hoạt EGA và sự phát triển bình thường của phôi sau này. (Nguồn: Pardinas và cs. 2026)
Kiêng xuất tinh kéo dài làm tăng sự tích lũy tổn thương DNA do oxy hóa (Gunes và cs. 2018). Việc khuyến khích xuất tinh thường xuyên hoặc sử dụng mẫu tinh trùng với thời gian kiêng ngắn (1 ngày) có thể giúp cải thiện chất lượng DNA tinh trùng trước khi thực hiện các kỹ thuật hỗ trợ sinh sản (Esteves 2025).
Sử dụng tinh trùng từ mô tinh hoàn (Testicular Sperm Extraction -TESE) ở những bệnh nhân có SDF cao trong tinh dịch xuất tinh (Esteves 2025). Tinh trùng tinh hoàn tránh được tác động của ROS tại mào tinh, giúp giảm tỷ lệ đứt gãy DNA tới 80% so với tinh trùng xuất tinh (Esteves 2025). Các nghiên cứu đã chứng minh chiến lược này cải thiện đáng kể tỷ lệ phôi sống sót và tỷ lệ trẻ sinh sống (Juan và cs. 2023, Esteves 2025). Tuy nhiên, đây vẫn là một quan điểm thực hành còn nhiều tranh luận vì tính chất xâm lấn.
Chọn lọc tinh trùng cho IVF/ICSI bằng phương pháp microfluidics hay hyaluronan cho phép chọn lọc những tinh trùng có độ di động tiến tới tốt và chỉ số SDF thấp nhất bằng cách mô phỏng cách thức chọn lọc tự nhiên trong đường sinh dục nữ (Juan và cs. 2023, Balder và cs. 2024, Esteves 2025).
Sử dụng các liệu pháp chống oxy hóa đường uống như vitamin C, vitamin E, selenium, và coenzyme Q10 có thể giúp giảm SDF ở nam giới gặp tình trạng stress oxy hóa (Esteves 2025). Tuy nhiên, cần thận trọng để tránh gây ra hiện tượng Reductive Stress (Stress khử), điều này có thể làm phá vỡ các cầu nối disulfide cần thiết cho sự nén chặt ổn định của chất nhiễm sắc (Esteves 2025, Aitken và cs. 2023).
Mặc dù cơ chế sửa chữa DNA đã được làm rõ ở mức độ phân tử, việc áp dụng vào lâm sàng vẫn là khoảng cách lớn bởi nhiều lý do:
(1) Các xét nghiệm SDF hiện nay chỉ cho biết số lượng tổn thương mà không thể dự báo liệu noãn có đủ năng lực để sửa chữa những lỗi cụ thể đó hay không.
(2) Phần lớn dữ liệu về sự sửa chữa này đều đến từ mô hình chuột, vốn dĩ quá trình EGA diễn ra sớm hơn (giai đoạn 2 tế bào) so với ở người (giai đoạn 8 tế bào), điều này dẫn đến những câu hỏi về loại suy kiến thức trên mô hình động vật và tiên lượng lâm sàng.
(3) Việc điều trị vô sinh chưa thật sự có sự liên kết giữa nam khoa và sản khoa, dẫn đến việc bỏ qua những dữ liệu nghiên cứu xâu chuỗi giữa chất lượng tinh trùng, khả năng sửa chữa của noãn, kết quả HTSS và sức khỏe trẻ sinh sống.
Tiếp tục làm rõ vai trò của các protein hỗ trợ sửa chữa mới được phát hiện như XRCC4-Like Factor (XLF) và Paralog of XRCC4 and XLF (PAXX). Đây là những thành phần thiết yếu trong con đường NHEJ, đóng vai trò quan trọng trong việc gắn nối các đầu đứt gãy DNA. Việc hiểu rõ chức năng của các protein này sẽ cho phép phát triển các xét nghiệm tiên lượng chính xác hơn về khả năng sống sót và tính toàn vẹn của hợp tử (Li và cs. 2025).
Nhằm khắc phục sự suy giảm mRNA và protein sửa chữa ở phụ nữ lớn tuổi, các nghiên cứu đề xuất sử dụng hoạt chất phục hồi năng lượng tế bào như Nicotinamide Adenine Dinucleotide (NAD+) (Newman và cs. 2022). Đồng thời, liệu pháp thay thế ty thể (Mitochondrial Replacement Therapy) đang được nghiên cứu để khôi phục trạng thái khỏe mạnh và năng lực sửa sai của noãn, mặc dù cần thêm các dữ liệu lâm sàng để tối ưu hóa hiệu quả (Pardiñas và cs. 2026).
Phát triển công nghệ Nano nhắm trúng đích thay vì các can thiệp triệu chứng hiện nay, sử dụng các hạt nano chức năng hóa được gắn các peptide đặc hiệu để đưa thuốc chống oxy hóa hoặc các tác nhân sửa chữa gen trực tiếp vào biểu mô tinh hoàn. Phương pháp này hứa hẹn sẽ giảm chỉ số SDF ngay từ quá trình sinh tinh (Aitken và cs. 2024).
- Aitken RJ, Lewis SEM. DNA damage in testicular germ cells and spermatozoa. When and how is it induced? How should we measure it? What does it mean? Andrology. 2023;11(8), 1545-1557.
- Aitken RJ. Paternal age, de novo mutations, and offspring health? New directions for an ageing problem. Hum Reprod. 2024;39(12), 2645-2654.
- García-Rodríguez A, Gosálvez J, Agarwal A, và cs. DNA Damage and Repair in Human Reproductive Cells. Int J Mol Sci. 2019;20(1), 31.
- Balder P, Jones C, Coward K, và cs. Sperm chromatin: Evaluation, epigenetic signatures and relevance for embryo development and assisted reproductive technology outcomes. Eur J Cell Biol. 2024;103(3), 151429.
- Esteves SC. From Double Helix to Double Trouble: Sperm DNA Fragmentation Unveiled – A Reproductive Urologist Perspective. Int Braz J Urol. 2025;51(1), e20249924.
- Gunes S, Sertyel S. Sperm DNA Damage and Oocyte Repair Capability. A Clinician’s Guide to Sperm DNA and Chromatin Damage. 2018;18, 321-346.
- Alvarez JG, García-Peiró A, Barros A, và cs. Double strand DNA breaks in sperm: the bad guy in the crowd. J Assist Reprod Genet. 2023;40(4), 741-746.
- Li N, Wang H, zou S, và cs. Perspective in the Mechanisms for Repairing Sperm DNA Damage. Reprod Sci. 2025;32(1), 41-51.
- Newman H, Catt S, Vining B, và cs. DNA repair and response to sperm DNA damage in oocytes and embryos, and the potential consequences in ART: a systematic review. Mol Hum Reprod. 2022;28(1), gaab071.
- Pardiñas ML, de Celis C, Gil J, và cs. Oocyte-mediated repair of sperm DNA fragmentation: a critical determinant of embryo viability. Reprod Biomed Online. 2026;52(2), 10516.