Nội dung chính
Tổn thương DNA của tinh trùng là một yếu tố quan trọng ảnh hưởng đến kết cục sinh sản. Quá trình xử lý các tổn thương này không diễn ra tại một thời điểm duy nhất mà trải dài từ giai đoạn sinh tinh cho đến sau khi thụ tinh, với sự tham gia của cả tế bào mầm nam và noãn. Tuy nhiên, khả năng sửa chữa thay đổi đáng kể theo từng giai đoạn phát triển của tinh trùng.
2.1 Các cơ chế sửa chữa ở tế bào mầm sinh tinh
Trong giai đoạn sinh tinh, các tế bào mầm sinh tinh có đầy đủ các cơ chế sửa chữa tổn thương DNA mạch đơn và mạch đôi, nổi bật trong số đó gồm (Hình 1) [1]:
Các cơ chế sửa chữa đứt gãy mạch đơn như:
- Sửa chữa sai cặp (Mismatch Repair - MMR): nhận diện các sai lệch nucleotide trên sợi DNA con thông qua phức hợp MSH2/MSH6, sau đó loại bỏ đoạn DNA bị lỗi nhờ exonuclease (EXO1) và tổng hợp lại đoạn đúng.
- Cắt bỏ nucleotide (Nucleotide Excision Repair - NER): loại bỏ các tổn thương DNA kích thước lớn bằng cách cắt bỏ đoạn DNA bị hư hại nhờ các protein XPA, XPC, XPG và ERCC1, sau đó được DNA polymerase và ligase nối lại.
- Cắt bỏ base (Base Excision Repair - BER): sửa chữa các tổn thương base, đặc biệt do stress oxy hóa, thông qua việc loại bỏ base hư bởi DNA glycosylase, sau đó tổng hợp lại DNA nhờ POLβ kết hợp với XRCC1 và ligase.
Đối với đứt gãy mạch đôi, tế bào chủ yếu sử dụng:
- Tái tổ hợp tương đồng (Homologous Recombination - HR): nhận diện đứt gãy thông qua các kinase cảm biến tổn thương DNA như ATM/ATR, xử lý đầu DNA nhờ phức hợp protein MRN và sử dụng trình tự DNA tương đồng làm khuôn để sửa chữa chính xác thông qua RAD51 và BRCA1/2.
- Nối đầu không tương đồng (Non-Homologous End Joining - NHEJ): sử dụng phức hợp Ku70/Ku80 và các protein như DNA-PKcs, XRCC4, LIG4 để nối trực tiếp hai đầu DNA mà không cần khuôn, do đó nhanh nhưng dễ gây sai sót.
- Nối đầu thay thế (Alternative End Joining - aEJ): được kích hoạt khi NHEJ bị rối loạn, sử dụng PARP1 và XRCC1/LIG1/3 để nối DNA, nhưng có độ chính xác thấp hơn.
- Bắt cặp sợi đơn (Single Strand Annealing - SSA): sửa chữa các đứt gãy có vùng tương đồng dài bằng cách cắt bỏ đoạn DNA và bắt cặp trực tiếp các vùng tương đồng, phụ thuộc vào phức hợp MRN và CtIP.
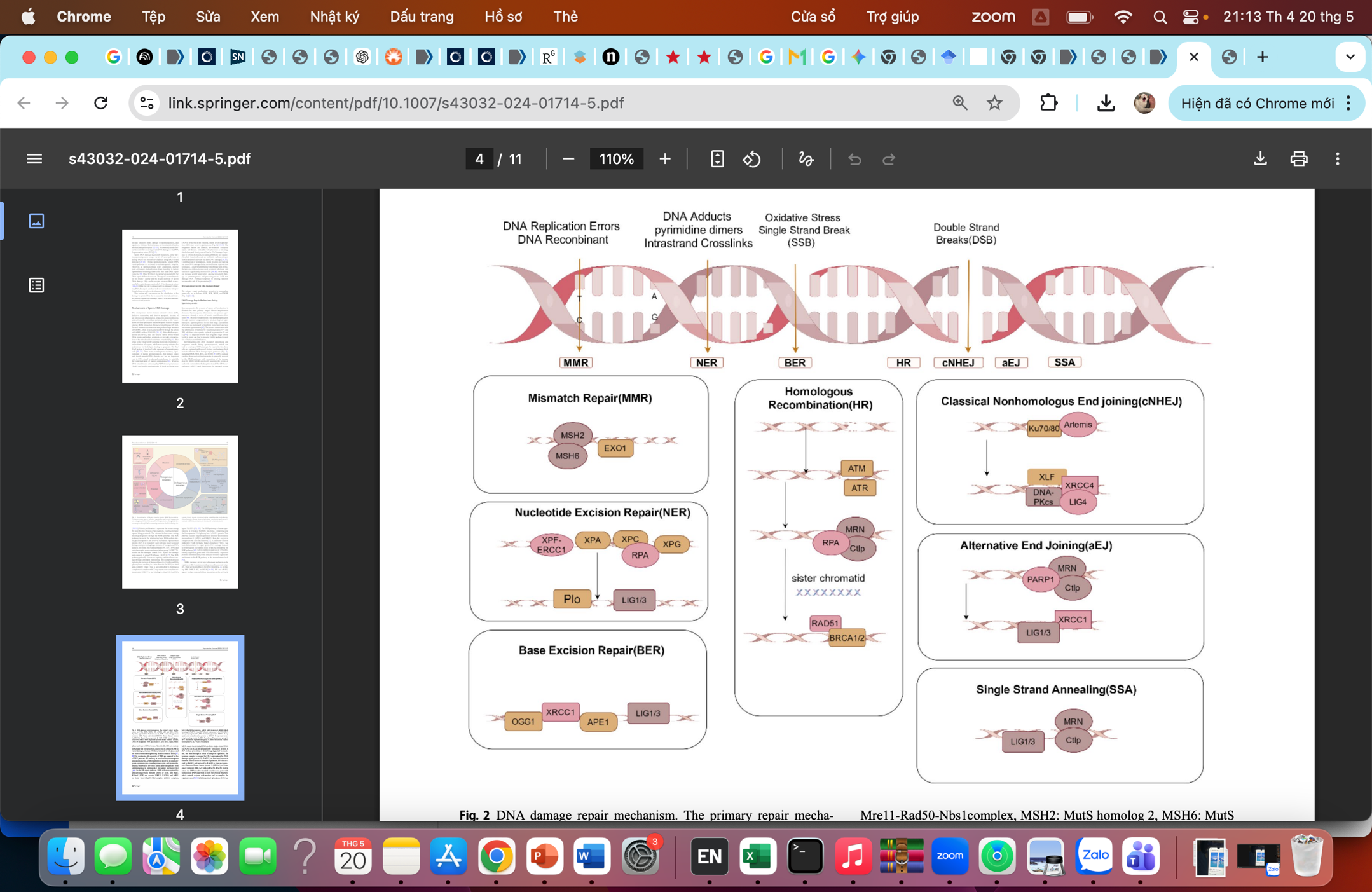
Hình 1: Các cơ chế sửa chữa DNA phổ biến [1]
2.2 Sự suy giảm năng lực sửa chữa trong giai đoạn biệt hoá tinh trùng trưởng thành
Các cơ chế sửa chữa DNA nêu trên hoạt động hiệu quả trong các giai đoạn nguyên phân và giảm phân, đảm bảo phần lớn tổn thương DNA được sửa chữa trước khi tinh trùng trưởng thành. Tuy nhiên, trong giai đoạn biệt hoá thành tinh trùng trưởng thành, sự thay thế histone bằng protamine làm nhiễm sắc chất cô đặc mạnh, khiến DNA gần như bất hoạt và hạn chế sự tiếp cận của các enzyme sửa chữa [2]. Đồng thời, việc tinh trùng đào thải hầu hết bào tương làm mất đi các bào quan và bộ máy dịch mã, điều dẫn đến khả năng sửa chữa DNA ở tinh trùng trưởng thành gần như mất đi hoàn toàn [3]. Ở giai đoạn này, đa số các cơ chế sửa chữa đều bị rút gọn tới mức tối thiểu, chỉ còn sót lại một lượng nhỏ enzyme nhận diện tổn thương bám lại trong nhân, điển hình là enzyme 8-oxoguanine DNA glycosylase-1 (OGG1) có khả năng nhận diện và cắt bỏ các base bị oxy hóa như 8-OHdG. Tuy nhiên, tinh trùng trưởng thành thiếu các protein downstream thiết yếu như APE1 và XRCC1, vốn cần thiết để xử lý vị trí mất base và hoàn tất quá trình sửa chữa [4]. Do đó, mặc dù có thể nhận diện tổn thương, tinh trùng không có khả năng tự hoàn tất quá trình sửa chữa DNA.
2.3 Cơ chế bảo vệ thụ động ở tinh trùng trưởng thành
Mặt khác, tinh trùng trong giai đoạn trưởng thành chuyển sang chiến lược bảo tồn DNA thông qua các cơ chế bảo vệ thụ động. Quan trọng nhất là sự nén chặt nhiễm sắc chất nhờ protamine, giúp DNA được đóng gói ở mức độ cao và giảm khả năng tiếp cận của các tác nhân gây tổn thương. Cấu trúc này tiếp tục được ổn định bởi các liên kết disulfide hình thành trong quá trình trưởng thành ở mào tinh, làm tăng độ bền vững của bộ gen [3]. Bên cạnh đó, do khả năng chống oxy hóa nội bào hạn chế, tinh trùng phụ thuộc chủ yếu vào hệ thống chống oxy hóa ngoại bào từ dịch mào tinh và tinh dịch, nơi cung cấp các yếu tố giúp trung hòa các loại oxy phản ứng như superoxide dismutase, catalase, glutathione peroxidase, glutathione, vitamin C, vitamin E và kẽm [3].
3.1 Vai trò của noãn
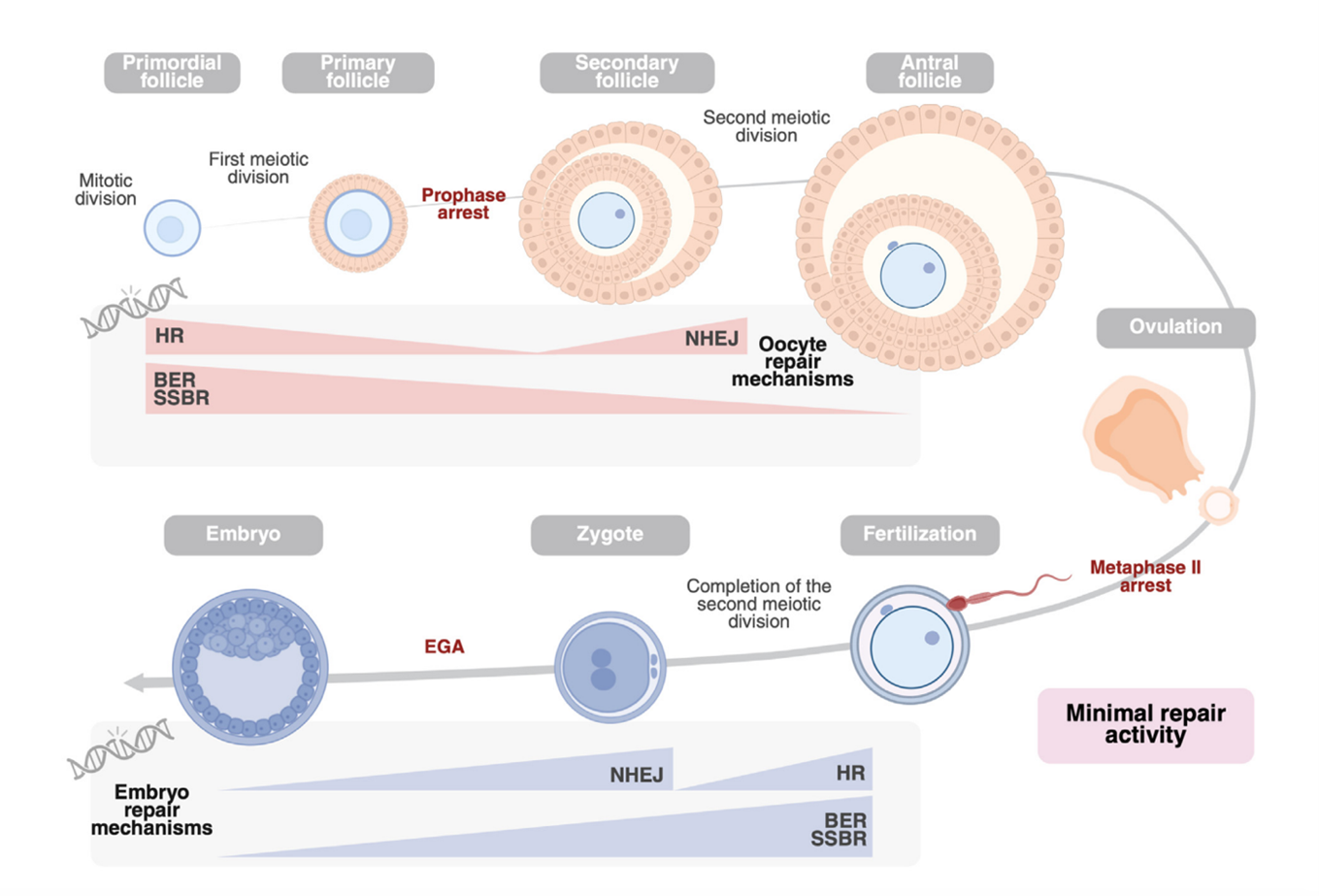
Sau khi thụ tinh, DNA của tinh trùng được giải nén và thay thế protamine bằng histone, cho phép các yếu tố sửa chữa từ noãn tiếp cận [5]. Quá trình sửa chữa diễn ra chủ yếu ở giai đoạn tiền nhân và trong pha tổng hợp DNA đầu tiên, tạo nên một “cửa sổ” quan trọng quyết định hiệu quả sửa chữa. Noãn sử dụng các cơ chế sửa chữa đứt gãy mạch đơn và BER để chủ yếu sửa các tổn thương nhẹ, trong khi HR và NHEJ sửa các đứt gãy mạch đôi. Việc lựa chọn cơ chế phụ thuộc vào chu kỳ tế bào: ở giai đoạn sớm sau thụ tinh khi DNA chưa được nhân đôi (pha G1), hợp tử không có khuôn tương đồng nên các tổn thương chủ yếu được sửa bằng NHEJ – cơ chế nhanh nhưng dễ gây sai sót; trong khi HR chỉ có thể hoạt động ở pha S/G2 khi đã có chromatid chị em làm khuôn [5, 6].
Đồng thời, hợp tử có thể điều chỉnh tiến trình chu kỳ tế bào nhằm tối ưu hóa quá trình sửa chữa DNA. Khi phát hiện tổn thương, các checkpoint như S-phase hoặc G2/M có thể được kích hoạt, làm chậm quá trình nhân đôi DNA, đặc biệt ở tiền nhân đực, qua đó kéo dài thời gian cho các cơ chế sửa chữa hoạt động trước khi bước vào phân bào [7]. Sau khi quá trình sửa chữa hoàn tất, các tín hiệu đáp ứng tổn thương DNA được tắt thông qua các cơ chế điều hòa, trong đó có vai trò của phosphatase WIP1 trong việc khử phosphoryl hóa các marker như γH2AX và p53, giúp tế bào tiếp tục chu kỳ phát triển bình thường [1, 5].
-
- Các yếu tố ảnh hưởng hiệu quả sửa chữa của noãn
Trong giai đoạn này, hiệu quả sửa chữa phụ thuộc vào hai yếu tố chính là chất lượng noãn và mức độ tổn thương DNA. Khi tuổi mẹ tăng, chất lượng noãn suy giảm kèm theo giảm biểu hiện các gen sửa chữa DNA, đặc biệt các gen liên quan đến sửa chữa đứt gãy mạch đôi như BRCA1, RAD51, MRE11 và ATM [8]. Đồng thời, sự lão hóa ty thể làm tăng stress oxy hóa do rối loạn chức năng ty thể và tích lũy các gốc oxy hóa phản ứng, cùng với giảm biểu hiện các enzyme chống oxy hóa tại ty thể như TXN2, SOD1 và PRDX3, khiến noãn trở nên dễ bị tổn thương hơn và suy giảm khả năng sửa chữa DNA [5]. Bên cạnh đó, các bệnh lý nền của người mẹ như béo phì hoặc hội chứng buồng trứng đa nang cũng có thể làm suy giảm thêm năng lực sửa chữa DNA của noãn [1]. Trong khi đó, mức độ tổn thương DNA tinh trùng khó được đánh giá chính xác ở cấp độ tinh trùng đơn lẻ, do các chỉ số phân mảnh DNA thường chỉ phản ánh giá trị trung bình của toàn bộ quần thể, làm phức tạp thêm việc dự đoán hiệu quả sửa chữa sau thụ tinh.
Sự chuyển giao hoạt hóa bộ gen phôi (Embryonic Genome Activation - EGA) diễn ra ở giai đoạn phôi 4-8 tế bào, khi phôi ngừng phụ thuộc vào nguồn mRNA của mẹ và tự kích hoạt bộ máy di truyền riêng. Tuy nhiên, từ giai đoạn này cho đến khi hình thành phôi nang, mức độ biểu hiện của nhiều gen sửa chữa cốt lõi (như OGG1, RAD50) và các miRNA điều hòa lại sụt giảm đáng kể so với noãn ban đầu. Khả năng sửa chữa suy giảm cùng xu hướng tăng cường sử dụng cơ chế nối đầu không tương đồng (NHEJ) làm tăng mạnh nguy cơ cố định các sai sót vào bộ gen [9]. Lúc này, các đứt gãy DNA không được xử lý sớm sẽ đẩy mức biểu hiện của protein cảnh báo p53 lên cao, đồng thời làm giảm RAD51 (protein chủ chốt của cơ chế sửa chữa chính xác HR) [1, 10]. Chuỗi tín hiệu bất lợi này khiến phôi mang tổn thương thường bị chậm phát triển, phân chia hỗn loạn hoặc kích hoạt apoptosis rõ rệt nhất ở các giai đoạn phôi muộn [7]. Dù rõ ràng kém hiệu quả hơn so với trước EGA, mạng lưới sửa chữa DNA chi tiết ở giai đoạn tiền làm tổ muộn này vẫn là một thách thức cần được nghiên cứu thêm.
Quá trình xử lý tổn thương DNA tinh trùng diễn ra liên tục từ giai đoạn sinh tinh đến phát triển phôi sớm, với sự chuyển giao dần năng lực sửa chữa từ tế bào mầm sinh tinh sang noãn và phôi. Trong khi tế bào mầm có đầy đủ cơ chế sửa chữa, tinh trùng trưởng thành chủ yếu dựa vào các cơ chế bảo vệ thụ động và phụ thuộc vào khả năng sửa chữa của noãn sau thụ tinh. Tuy nhiên, hiệu quả của quá trình này chịu ảnh hưởng bởi chất lượng noãn, mức độ tổn thương DNA tinh trùng và giai đoạn phát triển phôi, đặc biệt sau EGA khi khả năng sửa chữa có xu hướng suy giảm. Do đó, cần có thêm các nghiên cứu trong tương lai nhằm làm rõ giới hạn sửa chữa của noãn, cơ chế hoạt động sau EGA và ảnh hưởng lâu dài của các tổn thương DNA không được sửa chữa hoàn toàn đối với kết cục phôi và thế hệ sau.
- Li N, Wang H, Zou S, Yu X, Li J. Perspective in the mechanisms for repairing sperm DNA damage. Reprod Sci. 2025;32(1):41–51.
- Aitken RJ, Lewis SE. DNA damage in testicular germ cells and spermatozoa: when and how is it induced? How should we measure it? What does it mean? Andrology. 2023;11(8):1545–57.
- Aitken RJ, De Iuliis GN. On the possible origins of DNA damage in human spermatozoa. Mol Hum Reprod. 2009;16(1):3–13.
- Smith TB, Dun MD, Smith ND, Curry BJ, Connaughton HS, et al. The presence of a truncated base excision repair pathway in human spermatozoa that is mediated by OGG1. J Cell Sci. 2013;126(Pt 6):1488–97.
- Pardiñas ML, de Celis C, Gil J, Ortega-Jaen D, Martin A, Mercader A, et al. Oocyte-mediated repair of sperm DNA fragmentation: a critical determinant of embryo viability. Reprod Biomed Online. 2025:105165.
- Mao Z, Bozzella M, Seluanov A, Gorbunova V. DNA repair by nonhomologous end joining and homologous recombination during cell cycle in human cells. Cell Cycle. 2008;7:2902–6.
- Gawecka JE, Marh J, Ortega M, Yamauchi Y, Ward MA, Ward WS. Mouse zygotes respond to severe sperm DNA damage by delaying paternal DNA replication and embryonic development. PLoS One. 2013;8:e56385.
- Titus S, Li F, Stobezki R, Akula K, Unsal E, Jeong K, et al. Impairment of BRCA1-related DNA double-strand break repair leads to ovarian aging in mice and humans. Sci Transl Med. 2013;5.
- Jaroudi S, Kakourou G, Cawood S, Doshi A, Ranieri DM, Serhal P, et al. Expression profiling of DNA repair genes in human oocytes and blastocysts using microarrays. Hum Reprod. 2009;24(10):2649–55.
- Chen H, Liao SB, Cheung MP, Chow PH, Cheung AL, et al. Effects of sperm DNA damage on the levels of RAD51 and p53 proteins in zygotes and 2-cell embryos sired by golden hamsters without the major accessory sex glands. Free Radic Biol Med. 2012;53(4):885–92.